Elemental Phytoplankton Profiles

For many years, oceanographers have considered the Redfield ratio (C:N:P = 106:16:1) a cornerstone concept; it represents an emergent property that integrates biogeochemical cycles on basin scales over tens of thousands of years. The ratios hold very well for nitrate and phosphate in ocean waters and are thought to indicate healthy phytoplankton growth rates. When N:P ratios of less than 16 are measured, the findings are taken to indicate N limitation, whereas higher N:P ratios indicate P limitation. Since these ratios represent the net effect of biological activity in the system, marked deviations from the ratios may be expected in individual types of phytoplankton.
We measured the elemental composition of many phytoplankton grown under identical conditions (Quigg et al 2003; Ho et al 2003). Not only did we analyze C, N, and P, but also the major cations (K, Mg, Ca, Sr, etc...) and important trace elements (Fe, Mn, Cd, Co, Cu, Zn, etc…). Our findings can be examined in several ways:
(1) as an extended Redfield ratio which like the Redfield ratio, can be used to understand the biology of marine systems and be used in models of biogeochemical cycles. This is presented in Ho et al. (2003) Journal of Phycology
(2) to examine elemental signatures of different species, taxa and superfamilies in order to examine if patterns can be found which may elucidate forcing factors in the evolution and radiation of phytoplankton over the eons.
For an examination of the role of C, N and P, look at Martin et al. (2008) Palaeogeography, Palaeoclimatology, Palaeoecology.
For an examination of the role of trace elements (Fe, Mn, Zn, Co, Cu, and Cd) in the green and red superfamilies (or lineages), look at Quigg et al. (2003) Nature.
An introductory chapter on Ecological stoichiometry: Trace elements, look at Quigg, A. 2008. Trace Elements. Eds Sven Erik Jørgensen and Brian D. Fath. In, Ecological Stoichiometry in the Encyclopedia of Ecology. Vol. [5] 3564-3573 pp. Oxford, Elsevier.
For a more recent examination of the role of trace elements in the green and red superfamilies (or lineages), cyanobacteria and other groups, look at Quigg et al. (2011) Proceedings of the Royal Society: Biological Sciences.
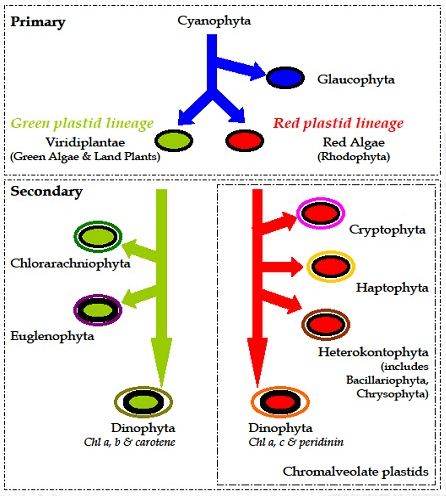
A single origin of plastids is hypothesized whereby a single-celled protist acquired and retained a free-living cyanobacterium. Over time, the cyanobacterium endosymbiont was reduced to a plastid and transmitted to subsequent generations. Three lineages arose from a primary engulfment event: the green plastid lineage (chl b-containing green algae and land plants, Viridiplantae), the red plastid lineage (chl c-containing red algae, Rhodophyta), and the Glaucocystophyta. Secondary endosymbiotic events occurred in both the green and red plastid lineages. In the green lineage, this gave rise to the Chlorarachniophyceae and Euglenophyceae. In the red plastid lineage, secondary endosymbiosis gave rise to the Chromalveolates which include the Cryptophyceae, the Heterokontophyta (such as the Bacillariophyceae and Chrysophyceae), the Haptophyceae, the chl c and peridinin containing Dinophyceae (but not the chl b containing dinoflagellates or those that arose as a result of a tertiary endosymbiotic event). (For more details; go to Quigg et al. 2011 Proceedings of the Royal Society: Biological Sciences).
We have now examined how the “Evolving Phytoplankton Stoichiometry Fueled Diversification of the Marine Biosphere”. For details, please read Martin, R. E. and Quigg, A. (2012) Geosciences. Special Issue on Paleontology and Geo/Biological Evolution.
Martin, R. E. and Quigg, A. 2013 Tiny Plants That Once Ruled the Seas. Scientific American. 308: 40-45.
Martin, R. E. and Quigg, A. 2012 Evolving Phytoplankton Stoichiometry Fueled Diversification of the Marine Biosphere. Geosciences. Special Issue on Paleontology and Geo/Biological Evolution. 2: 130-146.
Quigg, A., Irwin, A.J. and Finkel, Z.V. 2011 Evolutionary imprint of endosymbiosis of elemental stoichiometry: testing inheritance hypotheses. Proceedings of the Royal Society: Biological Sciences. 278: 526-534.
Finkel, Z. V., Beardall, J., Flynn, K. J., Quigg, A., Rees, T. A. K., and Raven, J. A. 2010 Phytoplankton in a changing world: cell size and elemental stoichiometry. Journal of Plankton Research,32: 119-137.
Beardall, J., Allen, D., Bragg, J., Finkel, Z. V., Flynn, K. J., Quigg, A., Rees, T. A. K., Richardson, A. and Raven, J. A. 2009 Allometry and stoichiometry of unicellular, colonial and multicellular phytoplankton. New Phytologist 181:295-309.
Quigg, A. 2008 Trace Elements. Eds Sven Erik Jørgensen and Brian D. Fath. In, Ecological Stoichiometry in the Encyclopedia of Ecology. Vol. [5] 3564-3573 pp. Oxford, Elsevier.
Martin, R. E., Quigg, A. and Podkovyrov, V. 2008 The evolution of ocean stoichiometry and diversification of the marine biosphere. Palaeogeography, Palaeoclimatology, Palaeoecology 258: 277–291.
Finkel, Z. V., Quigg, A., Chiampi, R., Schofield, O. and Falkowski, P. G. 2007 Phylogenetic diversity in Cd:P regulation by marine phytoplankton. Limnology and Oceanography, 52: 1131-1138.
Finkel, Z. V., Quigg, A., Raven, J. A., Reinfelder, J., Schofield, O. E. and Falkowski, P. G. 2006 Irradiance-induced changes in the elemental stoichiometry of marine phytoplankton. Limnology and Oceanography, 51: 2690-2701.
Falkowski, P. G., Katz, M. E., Knoll, A. H., Quigg, A., Raven, J. A., Schofield, O. and Taylor, F. J. R. 2004 The evolutionary history of eukaryotic phytoplankton. Science 305: 354-360.
Quigg, A., Finkel, Z.V., Irwin, A.J., Reinfelder, J.R., Rosenthal, Y., Ho, T-Y., Schofield, O., Morel, F. M. M. and Falkowski, P.G. 2003 The evolutionary inheritance of elemental stoichiometry in marine phytoplankton. Nature 425: 291-294.
The authors of this web page cordially invite contributions from all colleagues interested in building a data base that may be used in the future to further our understanding of ecological stoichiometry. Please submit as many details as possible about your particular phytoplankton(s), the assay system(s) and findings. Submissions should not be limited to multi-elemental studies, we would love findings on just N:P ratios, if you have additional information, such as DNA, RNA, quotas, etc…. and any publication details (or links). We hope to make this a community resource for students, postdoc’s, faculty, for modelers, to those that study molecules, the biosphere, and everything in between.
Amphidinium carterae
Gymnodinium chlorophorum

Pyramimonas parkeae

Ditylum brightwellii
Nannochloris atomus
Tetraselmis sp.
Dunaliella tertiolecta
Nitzschia brevirostris

Thalassiosira eccentrica

Emiliana huxleyi
Prorocentrum minimum

Thalassiosira weissflogii
Gephyrocapsa oceanica

Pycnococcus provasoli
Thoracosphaera heimii